1. Провідні шляхи пропріоцептивної (глибокої) чутливості. Складаються з пучків Голля і Бурдаха (рис. 502). З допомогою цих шляхів відбуваються рухи, які оцінюються свідомістю. Керованість рухів здійснюється за рахунок аферентних імпульсів з м'язів і суглобів рухомих частин тіла. Імпульси досягають постцентральна звивини кори тім'яної частки. Цей зворотний зв'язок забезпечує поступовість і координацію рухів. При пошкодженні шляхів пропріоцептивної чутливості хворий не може виконувати точні, співмірні, спритні рухи.
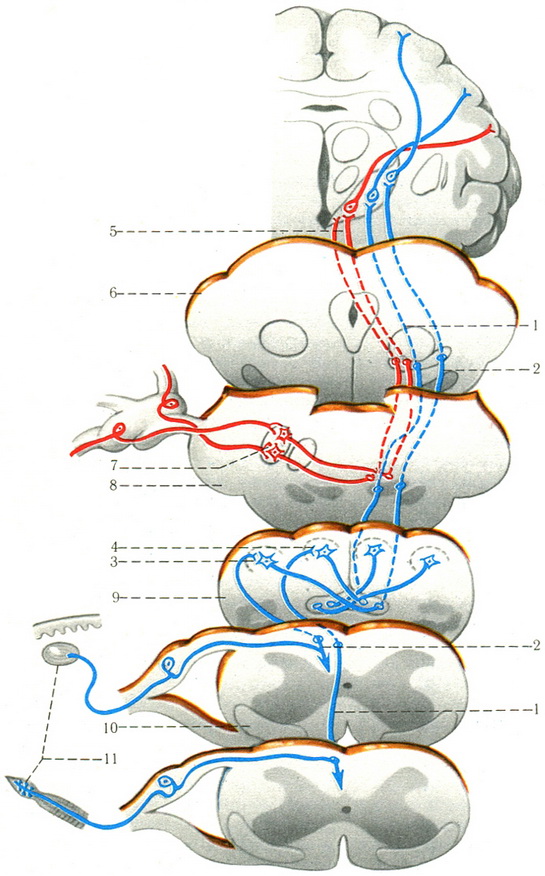
502. Схема пропріоцептивних провідних шляхів трійчастого нерва, Голля і Бурдаха (за Сентаготаі).
1 - шлях Голля; 2 - шлях Бурдаха; 3 - nucl. cuneatus; 4 - nucl. gracilis; 5 - чутливий шлях трійчастого нерва; 6 - середній мозок; 7-чутливе ядро V пари; 8 - міст; 9 - довгастий мозок; 10 - спинний мозок; 11 - проприорецептори шляхів Голля і Бурдаха.
Перші уніполярні чутливі нейрони шляхів Голля і Бурдаха розташовуються в спинномозкових вузлах (рис. 502). Їх рецептори - веретеноподібні тільця Кюне - починаються в м'язах, формуючи потім периферичний нерв. Аксони утворюють задній корінець, який набирає посегментно у біле речовина заднього канатика, об'єднуючись в тонкий (Голля) і клиновидний (Бурдаха) пучки. Тонкий пучок знаходиться ближче до медіальній борозні і складається з аксонів куприкових, крижових, поперекових, XII-VII грудних сегментів. Клиновидний пучок розташовується латеральніше тонкого пучка і об'єднує аксони від VIII - I грудних і VIII - I шийних сегментів.
Тонкий і клиновидний пучки закінчуються не на ядрах спинного мозку, а в тонкому і клиновидних ядрах довгастого мозку. Аксони клітин тонкого і клиновидного ядер (II нейрон) на кордоні з мостом утворюють медіальну петлю, яка контактує з клітинами вентролатерального ядра таламуса. З латеральної сторони до медіальної петлі приєднуються волокна спіноталамічного шляху. Аксони з ядер таламуса (III нейрон), пройшовши через задню частину внутрішньої капсули, закінчуються в корі верхньої тім'яної часточки (поля 5 і 7) і в передній центральній звивині (поля 4-6).
Частина волокон II нейронів пропріоцептивних чутливих шляхів прямує в мозочок через його нижні ніжки, беручи участь в механізмі координації рухів.
Існують проприоцептивные чутливі шляхи, які зв'язують ядра спинного мозку, довгастого мозку, моста, підкіркові утворення, экстрапирамидную підсистему з мозочком, які беруть участь у механізмах автоматичної координації рухів і тонусі м'язів, крім шляхів, замикаються в корі головного мозку. Ці механізми, як правило, проявляються при раптових порушення рівноваги або виконанні автоматичних рухів (ходьба, танці, лист та ін), виробляються в процесі вправ і під впливом соціальних моментів. Безусловнорефлекторные імпульси від всіх перерахованих вище утворень інтегруються в мозочок, який координує та визначає різні по точності руху. Імпульси з мозочка надають регулюючий гальмуючий вплив на ядра вестибулярного аналізатора і ретикулярної формації. Так як від вестибулярних ядер виникає преддверно-спинальний шлях, то за нього і ретикулоспинальному шляху настає пригнічення або полегшення функції альфа - і гамма-мотонейронів передніх стовпів спинного мозку і м'язових веретен рухових периферичних нервів. Таким чином, завдяки механізмам зворотного зв'язку через вестибулоспинальный і ретикулоспинальный шляху мозочок погоджує швидкі і повільні скорочення всіх м'язів. Мозочок нагадує блок регулювання, заснований на принципі зворотного зв'язку. Черв'як мозочка координує рухи при ходьбі і стоянні. У півкулі мозочка розташовані механізми дуже точної координації рухів, переважно для виконання рухів верхньої кінцівки. Черв'як підпорядкований корі мозочка, а вона функціонує під впливом кори головного мозку.